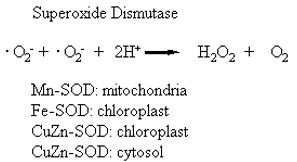
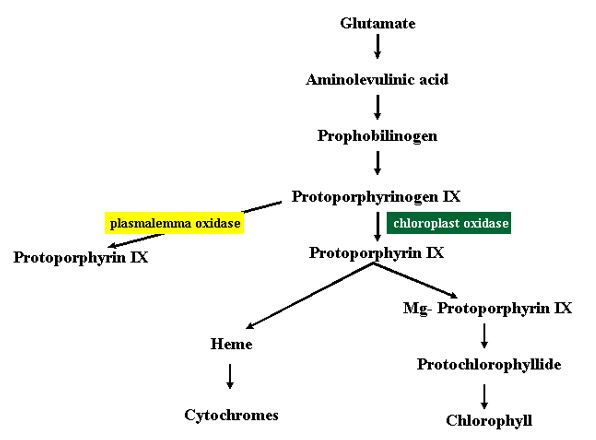
The presumed “spontaneous” oxidation of protoporphyrinogen IX to protoporphyrin IX is apparently accomplished by a protoporphyrinogen oxidising enzyme outside of the chloroplast, probably on the plasmalemma (Jacobs et al., 1991). This enzyme is presumably less sensitive than the chloroplast enzyme to the nitrodiphenyl ether inhibitors, and the presence of this second enzyme would explain many of the inconsistencies of the earlier data (Robeiz et al., 1990), the observed accumulation of protoporphyrin IX outside of the chloroplast, and the observed depletion of heme in the chloroplast. The major mechanism of action of acifluorfen and other diphenyl either herbicides is now thought to involve a blockage of the chloroplastic pathway shunting the protoporphyrinogen to an extra-chloroplastic pathway that first uncouples the regulation of the tetrapyrrole pathway on the chloroplast envelope by low heme levels, and secondly causes the accumulation of a photosensitising agent, protoporphrin IX in the cellular membranes.
The mode of action of the aminolevulinic acid-based herbicides is more clearly established because unlike other herbicides they were designed based on a knowledge of chlorophyll synthesis and free radical chemistry (Rebeiz et al., 1990). All formulations are based on the concept of supplying aminolevulinic acid to the plant as a precursor of tetrapyrrole synthesis (Fig. 15), in conjunction with a modulator that qualitatively and quantitatively alters the pattern of tetrapyrrole accumulation by selectively stimulating or inhibiting various steps in the biosynthetic pathway. The accumulated tetrapyrroles serve as photosensitizers that produce singlet oxygen in the light. The herbicide formulations are most effective if the sprayed plants remain in darkness for a few hours to allow tetrapyrrole accumulation before exposure to the light. After about 20 min illumination, the leaves develop isolated bleached areas that expand followed by wilting, desiccation and necrosis.
Although the photosensitising herbicides were assumed to be non-selective, laboratory and field trials have indicated that significant selectivity can be achieved by modifications of the formulations. This is caused by the different capacities of tissues and plants of accumulate tetrapyrroles and presumably to sequester or translocate the herbicides. Details of the mechanism of selectivity are given elsewhere (Rebeiz et al., 1990).
Some plants have apparently achieved increased resistance to acifluorfen by an enhanced scavenging capacity of oxygen free radicals (Gullner et al., 1991). Bean leaves treated with acifluorfen responded by higher levels of reduced glutathione and higher activity of glutathione reductase (Schmidt and Kunert, 1986). A tobacco line that was resistant to paraquat was also co-tolerant of acifluorfen (Gullner et al., 1991). This tolerance was associated with a greater inducibility of several components of the oxygen free radical scavenging system. Winter wheat (Triticum aestivum) seedlings that develop tolerance of freezing and of paraquat as they acclimate at 2°C, also develop increased resistance to acifluorfen (Bridger et al., 1994). Alfalfa plants that express a transgene of the Mn-SOD cDNA from Nicotiana have an increased LD50 for acifluorfen (McKersie et al., 1993).
SUMMARY AND CONCLUSIONS
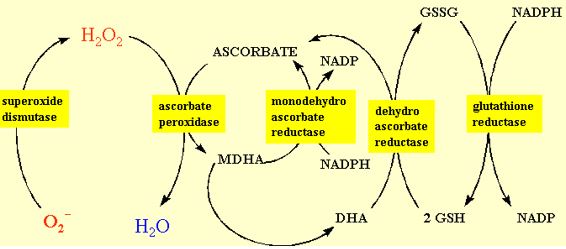
Oxygen free radicals or activated oxygen has been implicated in diverse environmental stresses in plants and animals and appears to be a common participation in most, if not all, degenerative conditions in eukaryotic cells. The peroxidation of lipids, the cross-linking and inactivation of proteins and mutations in DNA are typical consequences of free radicals, but because the reactions occur quickly and often are components of complex chain reactions, we usually can only detect their “footprints”. Activated forms of oxygen are important in the biosynthesis of “complex” organic molecules, in the polymerisation of cell wall constituents, in the detoxification of xenobiotic chemicals and in the defence against pathogens. Thus, the plant’s dilemma is not how to eliminate the activation of oxygen, but how to control and manage the potential reactions of activated oxygen. Complex systems of scavenging activated oxygen therefore exist in plant cells with complimentary and interdependent strategies. Some components such as the carotenoids prevent the formation of activated oxygen by competing for the energy leaked from the photosystems. Other components are lipid soluble and reside in the membrane bilayer to terminate the lipid peroxidation chain reactions. Still others, ascorbate and glutathione, are aqueous scavenger that detoxify activated oxygen directly or serve to recycle other protective components back to their reduced state. The enzymes that catalyse the synthesis, degradation and recycling of these antioxidants are essential to viability. Consequently, they are highly conserved among plants, and exist in multiple forms in different subcellular compartments and different tissues to allow precise regulation.
There are numerous sites of oxygen activation in the plant cell, which are highly controlled and tightly coupled to prevent release of intermediate products. Under stress situations, it is likely that this control or coupling breaks down and the process “dysfunctions” leaking activated oxygen. This is probably a common occurrence in plants especially when we consider that a plant has minimal mobility and control of its environment. These uncoupling events are not detrimental provided that they are short in duration and that the oxygen scavenging systems are able to detoxify the various forms of activated oxygen. If the production of activated oxygen exceeds the plant’s capacity to detoxify it, deleterious degenerative reactions occur, the typical symptoms being loss of osmotic responsiveness, wilting, and necrosis. At the subcellular level, disintegration of membranes and aggregation of proteins are typical symptoms. Therefore it is the balance between the production and the scavenging of activated oxygen that is critical to the maintenance of active growth and metabolism of the plant and overall environmental stress tolerance.
REFERENCES
Afanas’ev, I.B. 1985. Superoxide Ion: Chemistry and Biological Implications Volume 1. CRC Press, Boca Raton.
Alscher, R.G. 1989. Biosynthesis and antioxidant function of glutathione in plants. Physiol. Plant. 77:457-464.
Aono, M., Kubo, A., Saji, H., Natori, T., Tanaka, K. and Kondo, N. 1991. Resistance to active oxygen toxicity of transgenic Nicotiana tabacum that expresses the gene for glutathione reductase from Escherichia coli. Plant Cell Physiol. 32:691-697.
Asada, K. 1992. Ascorbate peroxidase – hydrogen peroxide-scavenging enzyme in plants. Physiol. Plant. 85: 235-241.
Bannister, J.V., Bannister, W.H. and Rotils, G. 1987. Aspects of the structure, function and applications of superoxide dismutase. CRC Crit. Rev. Biochem. 22:110-180.
Barber, J. and Andersson, B. 1992. Too much of a good thing: light can be bad for photosynthesis. Trends Biochem. Sci. 17:61-66.
Beaumont, F., Jouvc, H-M., Cagnan, J., Gillard, J. and Pelment, J. 1990. Purification and properties of a catalase from potato tubers (Solanum tuberosum). Plant Sci. 72:19-26.
Beyer, W., Imlay, J., Fridovich, I. 1991. Superoxide Dismutases. Prog. Nucl. Acid Res. 40:221-253.
Bielawski, W. and Joy, K.W. 1986. Reduced and oxidized glutathione and glutathione-reductase activity in tissues of Pisum sativum. Planta, 169:267-272.
Bowler, C., Slooten, L., Vandenbranden, S., De Rycke, R., Botterman, J., Sybesma, C., Van Montagu, M. and Inzé, D. 1991. Manganese superoxide dismutase can reduce cellular damage mediated by oxygen radicals in transgenic plants. EMBO. J. 10:1723-1732.
Bowler, C. and Van Montague, M. and Inzé, D. 1992. Superoxide dismutase and stress tolerance. Ann Rev. Plant Physiol. Plant Mol. Biol. 43:83-116.
Bowler C., Van Camp W. , Van Montagu M. and Inze D. 1994. Superoxide dismutase in plants. Critical Rev. Plant Sci. 13: 199-218
Bradley, D.E., Min. D.B. 1992. Singlet oxygen oxidation of foods. Cat. Rev. Food Sci. Nutri. 31: 211-236.
Bridger, G.M., Yang, W., Falk, D.E. and McKersie, B.D. 1994. Cold acclimation increases tolerance of activated oxygen in winter cereals. J. Plant Physiol. 144:235-240.
Brot, N. and Weissbach, H. 1982. The biochemistry of methionine sulfoxide residues in proteins. Trends Biochem. Sci. 7:137-139.
Burton, G.W. and Ingold, K.U. 1984. ß-carotene: an unusual type of lipid antioxidant. Science. 224:569-573.
Cakmak, I. and Marschner, H. 1988. Enhanced superoxide radical production in roots of zinc-deficient plants. J. Expt. Bot. 39:1449-1460.
Calderbank, A. 1968. The bipyridylium herbicides. Adv. in Pest Control Res. 8:127-135.
Chen, G.X. and Asada, K. 1989. Ascorbate peroxidase in tea leaves – occurrence of 2 isozymes and the differences in their enzymatic and molecular properties. Plant and Cell Physiology. 30:987-998.
Chichiricco, G., Ceru, M.P., D’Alessandro, A., Oratore, A., and Avigliano, L. 1989. Immunohistochemical localisation of ascorbate oxidase in Cucurbita pepo medullosa, Plant Sci. 64:61-66.
Creisson, G., Edwards, E.A., Anard, C. Wellburn A. and Mullineaux, P. 1992. Molecular characterization of glutathione reductase cDNAs from pea (Pisum sativum L. ). Plant J. 2:129-131.
Criqui, M.C., Jamet, E., Parmentier, Y., Marbach, J., Darr, A. and Fleck, J. 1992. Isolation and characterization of a plant cDNA showing homology to animal glutathione peroxidases. Plant Mol. Biol. 18: 623-627.
Davies, K.J.A. 1987. Protein damage and degradation by oxygen radicals. I General aspects. J. Biol. Chem. 162:9895-9901.
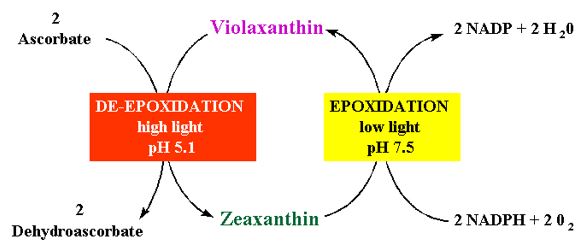
Demmig-Adams, B. and Adams, W.W. 1993. The Xanthophyll Cycle. In: R.G. Alscher and J.L. Hess eds. Antioxidants in Higher Plants. CRC Press, Baco Raton pp. 59-90.
Diplock, A.T., Machlin, L.J., Packer, L., and Pryor, W.A. 1989. Eds., Vitamin E: Biochemistry and Health Implications. Ann. N.Y. Acad. Sci. Vol. 570 p.555.
Dodge, J.D. and Lawes, G.B. 1974. Some effects of the herbicides diquat and morfamquat on the fine structure of leaf cells. Weed Res. 14:45-49.
Doke, N. and Ohashi, Y. 1988. Involvement of an O2-generating system in the induction of necrotic lesions on tobacco leaves infected with tobacco mosaic virus. Physiol. Mol. Plant. Pathol. 32:163-175.
Doke, N., Miura, Y., Chai, H-B, Kawakita, K. 1991. Involvement of Active Oxygen in induction of plant defense response against infection and injury. In: Active Oxygen/Oxidative Stress and Plant Metabolism. Pell E.J. and Steffen K.L. (eds) American Soc. Plant Physiol. Rockville, M.D. pp. 84-96.
Doke N., Miura Y., Sanchez L. M. and Kawakita K. 1994. Involvement of superoxide in signal transduction: responses to attack by pathogens, physical and chemical shocks, and UV irradiation. In: C. H. Foyer and P. M. Mullineaux (eds). Causes of photooxidative stress and amelioration of defense systems in plants. CRC Press Inc. Boca Raton, FL, USA. pp. 177-197
Drotar, A., Phelps, P. and Fall, R. 1985. Evidence for glutathione peroxidase activities in cultured plant cells. Plant Science 42:35-40.
Edwards, E.A., Rawsthorne, S. and Mullineaux, P.M. 1990. Subcellular distribution of multiple forms of glutathione reductase in leaves of pea (Pisum sativum L.). Planta 180:278-284.
Elstner, E.F. 1982. Oxygen activation and oxygen toxicity. Ann. Rev. Plant Physiol. 33:73-96.
Elstner, E.F. 1991. Mechanisms of oxygen activation in different compartments of plant cells In: Active oxygen/oxidative stress and plant metabolism. Pell E.J. and Steffen K.L. (eds) American Soc. Plant Physiol. Rockville, M.D. pp. 13-25.
Farr, S.B. and Kogoma, T. 1991. Oxidative stress responses in Escherichia coli and Salmonella typhimurium. Microbiol. Rev. 55:561-585.
Farrington, J.A., Ebert, H., Land, E.J. and Fletcher, K. 1973. Bipyridylium quaternary salts and related compounds. V. Pulse radiolysis studies of the reaction of paraquat radical with oxygen. Implications for mode of action of bipyridyl herbicides. Biochim. Biophys. Acta 314:372-381.
Feierabend, J., Schaan, C., and Hertwig, B. 1992. Photoinactivation of catalase occurs under both high- and low-temperature stress conditions and accompanies photoinhibition of photosystem II. Plant Physiol. 100:1554-1561.
Fenton, H.J.H. 1894. Oxidation of tartaric acid in the presence of iron. J. Chem. Soc. 65:899.
Fenton, H.J.H. 1899. Oxidation of certain organic acids in the presence of ferrous salts. Proc. Chem. Soc. 25:224.
Ferguson, D.L. and Burke, J.J. 1992. A new method of measuring protein-methionine-S-oxide reductase activity. Plant Physiol. 100:529-532.
Fita, I. and Rossmann, M.G. 1985. The active center of catalase. J. Mol. Biol. 185:21-37.
Foyer, C.H. and Halliwell, B. 1976. The presence of glutathione and glutathione reductase in chloroplasts: a proposed role in ascorbic acid metabolism. Planta. 133:21-25.
Foyer, C., 1993. Ascorbic acid. In: Antioxidants in Higher Plants. R.G. Alscher and J.L. Hess (eds) CRC Press, Boca Raton, pp.31-58.
Foyer, C. H., Descourvieres P. and Kunert K. J. 1994. Protection against oxygen radicals: an important defence mechanism studied in transgenic plants. Plant, Cell and Environment 17: 507-523
Frankel, E.N. 1985. Chemistry of free radical and singlet oxidation of lipids. Progress in Lipid Research. 23:197-221.
Fridovich, I. 1970. Quantitative aspects of the production of superoxide anion radical by milk xanthine oxidase. J. Biol Chem 245:4053-4057.
Fryer, M.J. 1992. The antioxidant effects of thylakoid vitamin E (a-tocopherol). Plant Cell Environ. 15:381-392.
Fuerst, E.P., Nakatani, H.Y., Dodge, A.D., Penner, D. and Arntzen, C.J. 1985. Paraquat resistance in Conyza. Plant Physiol. 77:984-989.
Fuerst, E.P. and Vaughn, K.C. 1990. Mechanisms of paraquat resistance. Weed Technol. 4:150-156.
Gardner, P.R. and Fridovich, I. 1991. Superoxide sensitivity of Escherichia coli 6-phosphogluconate dehydratose. J. Biol. Chem. 266:1478-1483.
Gebicki, J.M. and Bielski, B.H.J. 1981. Comparison of the capacities of the perhydroxyl and superoxide radicals to initiate chain oxidation of linoleic acid. J. Am. Chem. Soc. 103: 7020-7022.
Gressel, J. and Galun E. 1994. Genetic controls of photooxidant tolerance. In: C. H. Foyer and P. M. Mullineaux (eds). Causes of photooxidative stress and amelioration of defense systems in plants. CRC Press Inc. Boca Raton, FL, USA. pp. 237-273.
Gross, G.G., Janse, C. and Elstner, E.F. 1977. Involvement of malate, monophenols and superoxide radical in hydrogen peroxide formation by isolated cell walls from horseradish. (Armoracia lapathifolia Gilib.) Planta 136:271-276.
Gross, G.G. 1980. The biochemistry of lignification. Adv. Bot. Res. 8:25-63.
Gruber, M.Y., Glick, B.R. and Thompson, J.E. 1990. Cloned manganese superoxide dismutase reduces oxidative stress in Escherichia coli and Anacystis nidulans. Proc. Nat. Acad Sci. (U.S.A.) 87:2603-2612.
Gullner, G., Komives, T. and Kiroly, L. 1991. Enhanced inducibility of antioxidant systems in a Nicotiana tabacum L. biotype results in acifluorfen resistance. Naturforsch. 46c:875-881.
Haber, F. and Weiss, J. 1934. The catalytic decomposition of hydrogen peroxide by iron salts. Proc. Royal Soc. A. 147:332.
Harper, D.B., Harvey, B.M.R. 1978. Mechanism of paraquat tolerance in perennial ryegrass. II. Role of superoxide dismutase, catalase and peroxidase. Plant Cell Environ. 1:211-215.
Härtel, H., Haseloff, R.F., Ebert, B., Rank, B. 1992. Free radical formation in chloroplasts. Methyl viologen action. J. Photochem. Photobiol. B: Biol. 12: 375-387.
Hausladen, A. and Alscher, R.G. 1993. Glutathione In: Antioxidants in Higher Plants. R.G. Alscher and J.L. Hess (eds) CRC Press, Boca Raton. pp. 1-30.
Hell, R. and Bergmann, L. 1990. Glutamylcysteine synthetase in higher plants: catalytic properties and subcellular localization. Planta 180:603-612.
Hell, R. and Bergmann, L. 1988. Glutathione synthetase in tobacco suspension cultures: catalytic properties and localization. Physiol. Plant 72:70-76.
Hertwig, B., Steb, P. and Feierabend, J. 1992. Light dependence of catalase synthesis and degradation in leaves and the influence of interferring stress conditions. Plant Physiol. 100:1547-1553.
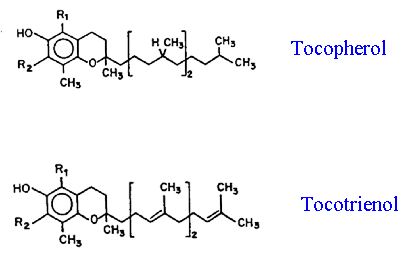
Hess, J.L. 1993. Vitamin E, a-tocopherol. In: Antioxidants in Higher Plants. R.G. Alscher and J.L. Hess (eds) CRC Press. Boca Raton, pp.111-134.
Hohn, D.C. and Lehere, R.L. 1975. NADPH oxidase deficiency in X-linked chronic granulomatous disease. J. Clin Invest. 53:707-713.
Imlay, J.A. and Linn, S. 1986. DNA damage and oxygen radical toxicity. Science 240:1302-1309.
Jacobs, J.M., Jacobs, N.J., Sherman, T.D. and Duke, S.O. 1991. Effect of diphenyl ether herbicides in oxidation of photoporphyrinogen to protoporphyrin in organellar and plasma membrane enriched fractions of barley. Plant Physiol. 97:197-203.
Johnson, W.O., Kollman, G.E., Swithenbank, C. and Yih, R.Y. 1978. RH-6201 (Blazer): A new broad spectrum herbicide for postemergence use in soybeans. J. Agric. Food Chem. 26:285-286.
Kelner, M.J. and Bagnell, R. 1990. Glutathione-dependent enzymes alone can produce paraquat resistance. Free Rad. Biol. Med. 9:149-153.
Kendall, E.J. and McKersie, B.D. 1989. Free radical and freezing injury to cell membranes of winter wheat. Physiol. Plant. 76:86-94.
Kirkman, H.N., Galiano, S. and Gaetani. 1987. The function of catalase-bound NADPH. J. Biol. Chem. 262:660-666.
Klapheck, S. 1988. Homoglutathione: isolation, quantification and occurrence in legumes. Physiol. Plant 74:727-732.
Kouiji, H., Masuda, T. and Matsunaka, S. 1989. Action mechanism of diphenyl ether herbicides: stimulation of 5-aminolevulinic acid synthesizing system activity. Pestic. Biochem. Physiol. 33:230-238.
Kuroda,H., Sagisaka, S., Asada, M. and Chiba, K. 1991. Peroxide-scavenging systems during cold acclimation of apple callus in culture. Plant and Cell Physiology. 32:635-641.
Lehnen, L.P. Jr., Sherman, T.D., Becerril, J.M. and Duke, S.O. 1990. Tissue and cellular localization of acifluorfen-induced porphyrins in cucumber cotyledons. Pestic. Biochem. Physiol 37:239-248.
Leshem, Y.Y., 1992. Plant Membranes: A Biophysical Approach to Structure, Development and Senescence. Kluwer Academic Publishers Dordrecht, The Netherlands, 266 p.
Lindqvist, Y., Branden, C.L., Mathews, F.S., and Lederer, F. 1991. Spinach glycolate oxidase and yeast flavocytochrome b2 are structurally homologous and evolutionarily related enzymes with distinctly different function and flavin mononucleotide binding. J. Biol. Chem. 266:3198-3207.
Loewus, F.A. 1988. Ascorbic acid and its metabolic products. In: The Biochemistry of Plants, Vol. 14. Preiss, J. (ed) Academic Press, New York, p. 85-107.
Loschen, G., Azzi, A. and Floheßp, L. 1973. Mitochondrial H2O2 formation: Relationship with energy conversion. FEBS Lett 33:84-88.
Loschen, G., Azzi, A., Richter, C. and Floheßp, L. 1974. Superoxide radicals as precursors of mitochondrial hydrogen peroxide. FEBS Lett 42:68-72.
Malan, C., Greyling, M.M. and Gressel, J. 1990. Correlation between Cu/Zn superoxide dismutase and glutathione reductase, and environmental and xenobiotic stress tolerance in maize inbreds. Plant Sci. 69:157-166.
Mann, T., and Kleilin, D. 1938. Homocuprein and heptacuprein, copper-protein compounds of blood and liver in mammals. Proc. R. Soc. London B. 126:303-315.
Marx, J.L. 1985. Oxygen free radicals linked to many diseases. Science. 235-529-531.
Mathis, P. and Kleo, J. 1973. The triplet state of ß-carotene and of analog polyenes of different length. Photochem. Photobiol. 18:343-346.
Matringe, M., Camadro, J-M., Labbe, P. and Scalla, R. 1989. Protoporphyrinogen oxidase as a molecular target for diphenyl ether herbicides. Biochem J. 260:231-235.
Matsumoto, H. and Duke, S.O. 1990. Acifluorfen-methyl effects on porphyrin synthesis in Lemna pousicostata. Hegelm. 6746. J. Agr. Food Chem. 38:2066-2071.
Mayasich, J.M., Nandihalli, U.B., Liebl, R.A. and Rebeiz, C.A. 1990. The primary mode of action of acifluorfen-Na in intact seedligns is not via tetrapyrrole accumulation during the first dark period following treatment. Pestic. Biochem. Physiol. 36:259-268.
McCord, J.M. and Fridovich, I. 1969. Superoxide dismutase, an enzymatic function for erythrocuprein. J. Biol. Chem. 244:6049-6055.
McKersie, B.D., Hoekstra, F., and Krieg, L. 1990. Differences in the susceptibility of plant membrane lipids to peroxidation. Biochim. Biophys. Acta. 1030:119-126.
McKersie, B.D., Chen, Y., de Beus, M., Bowley, S.R., Bowler, C., Inzé, D., D’Halluin, K. and Boterman, J. 1993. Superoxide dismutase enhances toleance of freezing stress in transgenic alfalfa (Medicago sativa L.). Plant Physiol. (in press).
Mees, G.C. 1960. Experiments on the herbicidal action of 1,l’-ethylene-2,2′-dipyridylium dibromide. Ann. Appl. Biol. 48:601-612.
Meister, A. 1988. Glutathione metabolism and its selective modification. J. Biol. Chem. 263:17205-17208.
Morré, D.J., Brightman, A.O., Wu, L.Y., Barr, R. Leak, B. and Crane, F.L. 1988. Role of plasma membrane redox activities in elongation growth in plants. Physiol. Plant 73: 187-193.
Oleinick, N.L., Chiu, S., Ramakrishman N. and Xue, L. 1986. The formation, identification, and significance of DNA-protein cross-links in mammalian cells. Brit. J. Cancer 55: Suppl. 8:135-140.
Pallett, K.E, and Young, A.J. 1993. Caratenoids. pp. 91-110. In: Eds. R.G. Alscher and J.L. Hess. Antioxidants in Higher Plants. CRC Press, Boca Raton.
Polle, A., Chakrabarti, K., Schürmann, W., and Rennenberg, H. 1990. Composition and properties of hydrogen peroxide decomposing systems in extracellular and total extracts from needles of Norway spruce (Picea abies L., karst). Plant Physiol. 94:312-319.
Preston, C., Holtum, J.A.M. and Powles, S.B. 1992. On the mechanism of resistance to paraquat in Hordeum glaucum and H. leporium. Plant Physiol. 100:630-636.
Price, A., Lucas, P.W. and Lea, P.J. 1990. Age dependent damage and glutathione metabolism in ozone fumigated barley: a leaf section approach. J. Exptl. Bot. 41:1309-1317.
Rebeiz, C.A., Reddy, K.N., Nandihalli, U.B. and Velu, J. 1990. Tetrapyrole-dependent photosythetic herbicides. Photochem. Photobiol. 52:1099-1117.
Redinbaugh, M.G., Wadsworth, G.J. and Scandalias, J.G. 1988. Characterization of catalase transcripts and their differential expression in maize. Biochim. Biophys. Acta. 951:104-116.
Rennenberg, H. 1982. Glutathione metabolism and possible biological roles in higher plants. Phytochem. 21:2771-2781.
Rich, P.R., Bonner, W.D. Jr. 1978. The sites of superoxide anion generation in higher plant mitochondria. Arch. Biochem. Biophys. 188:206-213.
Rüegsegger, A., Schmutz, D. and Brunold, C. 1990. Regulation of glutathione biosynthesis by cadmium in Pisum sativum L. Plant Physiol. 93:1579-1584.
Sandalio, M. and Del Rio, L.A. 1988. Intraorganellar distribution of superoxide dismutase in plant peroxisomes (glyoxysomes and leaf peroxisomes). Plant Physiol. 88:1215-1218.
Scandalias, J.G. 1990. Response of plant antioxidant defense genes to environmental stress. Adv. Genet. 28:1-41.
Scandalias, J.G. 1993. Oxygen stress and superoxide dismutase. Plant Physiol. 101:7-12.
Schmidt. A., and Kunert, K.J. 1986. Lipid peroxidation in higher plants. The role of glutathione reductose. Plant Physiol 82:700-702.
Shaaltiel, Y. and Gressel, J. 1986. Multienzyme oxygen radical detoxifying system correlated with paraquat resistance in Conyza bonariensis. Pestic. Biochem. Physiol. 26:22-28.
Smith, I.K., Kendall, A.C., Keys, A.J., Turner, J.C. and Lea, P.J. 1985. The regulation of the biosynthesis of glutathione in leaves of barley (Hordeum vulgare L.). Plant Sci. 41:11-17.
Smith, I.K., Vierheller, T.L. and Thorne, C.A. 1989. Properties and functions of glutathione reductase in plants. Physiol. Plant. 77:449-456.
Smith, J. and Shrift, A. 1979. Phytogenetic distribution of glutathione peroxidase. Comp. Biochem. Physiol. 63B:39-44.
Stadtman, E.R. 1986. Oxidation of proteins by mixed-function oxidation systems: implication in protein turnover, aging and neutrophil function. Trends Biochem. Sci. 11:11-12.
Steinkamp, R. and Rennenberg, H. 1984. Glutamyltranspeptidase in tobacco suspension cultures: catalytic properties and subcellular localization. Physiol. Plant. 61:251-256.
Tepperman, J.M. and Dunsmuir, P. 1990. Transformed plants with elevated level of chloroplastic SOD are not more resistant to superoxide toxicity. Plant Mol. Biol. 14:501-511.
Timmerman, K.P. 1989. Molecular characterization of corn glutathione-S-transferase isozymes involved in herbicide detoxification. Physiol. Plant. 77:465-471.
Turrens, J.F., Freeman, B.A., Crapo, J.D. 1982. Hyperoxia increases H2O2 release by lung mitochondria and microsomes. Arch. Biochem. Biophys. 217:411-421.
Van Camp W., van Montagu M., Inze D. 1994. Superoxide dismutases. In: C. H. Foyer and P. M. Mullineaux (eds). Causes of photooxidative stress and amelioration of defense systems in plants. CRC Press Inc. Boca Raton; USA. 317-341
Vaughn, K.C. and Fuerst, E.P. 1985. Structural and physiological studies of paraquat-resistant Conyza. Pestic. Biochem. Physiol. 24:86-94.
Vianello, A., Macri, F. 1991. Generation of superoxide anion and hydrogen peroxide at surface of plant cells. J. Bioenerg. Biomemb. 23:409-423.
Wilson, D.O. and McDonald, M.B. Jr. 1986. The lipid peroxidation model of seed aging. Seed Sci. Tech. 14:269-300.
Winston, G.W., Cederbaum, A.I. 1983a. Oxyradical production by purified components of the liver microsomal mixed-function oxidase system I: Oxidation of hydroxyl radical scavenging agents. J. Biol Chem 258:1508-1513.
Witkowski, D.A. and Halling, B.P. 1989. Inhibition of plant protoporphyrinogen oxidase by the herbicide acifluorfen-methyl. Plant Physiol. 90:1239-1242.
Young, A.J. 1991. Inhibition of carotenoid biosyntheses. pp.131-171. In: Eds. N.R. Baker and M.P. Percival. Herbicides. Elsevier Publishers.
Young, A.J. 1991. The photoprotective role of carotenoids in higher plants. Physiol. Plant. 83:702-708.
Bryan D. McKersie
Dept of Crop Science, University of Guelph (December, 1996)